.png?width=1900&name=blog_banner%20(1).png "blog_banner (1)")
Forbrain is a user-friendly learning tool that uses a bone conductor and a series of dynamic filters to provide feedback to the user.
State-of-the-art
Forbrain is thought to improve perception by optimizing all the components of the audio-vocal loop. It is considered a product that can improve speech, fluency, memory, focus, coordination, and many other sensory functions, resulting in several adjustments in the psychological/cognitive/emotional domain.
The use of a bone conductor by Forbrain extends the possibilities of stimulus manipulation and presentation in an unprecedented way, resulting in new ways to enrich the acoustic environment. Through bone conduction (bone vibration), the traveling waveform of a sound reaches the inner ear, induces a traveling wave in the basilar membrane, and stimulates the cochlea by the same mechanisms as normal air conduction (Stenfelt et al., 2003). This way, the cochlea, its basilar membrane, and the encoding of the incoming sound signal into a neural pulse, create a bottleneck through which both air-conducted and bone-conducted sounds converge on their way to the central auditory system (Stenfelt et al., 2003; 2005). If the amplitude and phase of a particular sound are appropriately adjusted, its cochlear signal should be canceled out. Therefore, through the dynamic filter used in Forbrain, unexpected and random changes in the bone conduction signal are introduced that may eventually cancel out or distort the air conduction signal to the very same sounds, resulting in unexpected, rare, and potentially surprising changes in the auditory signal.
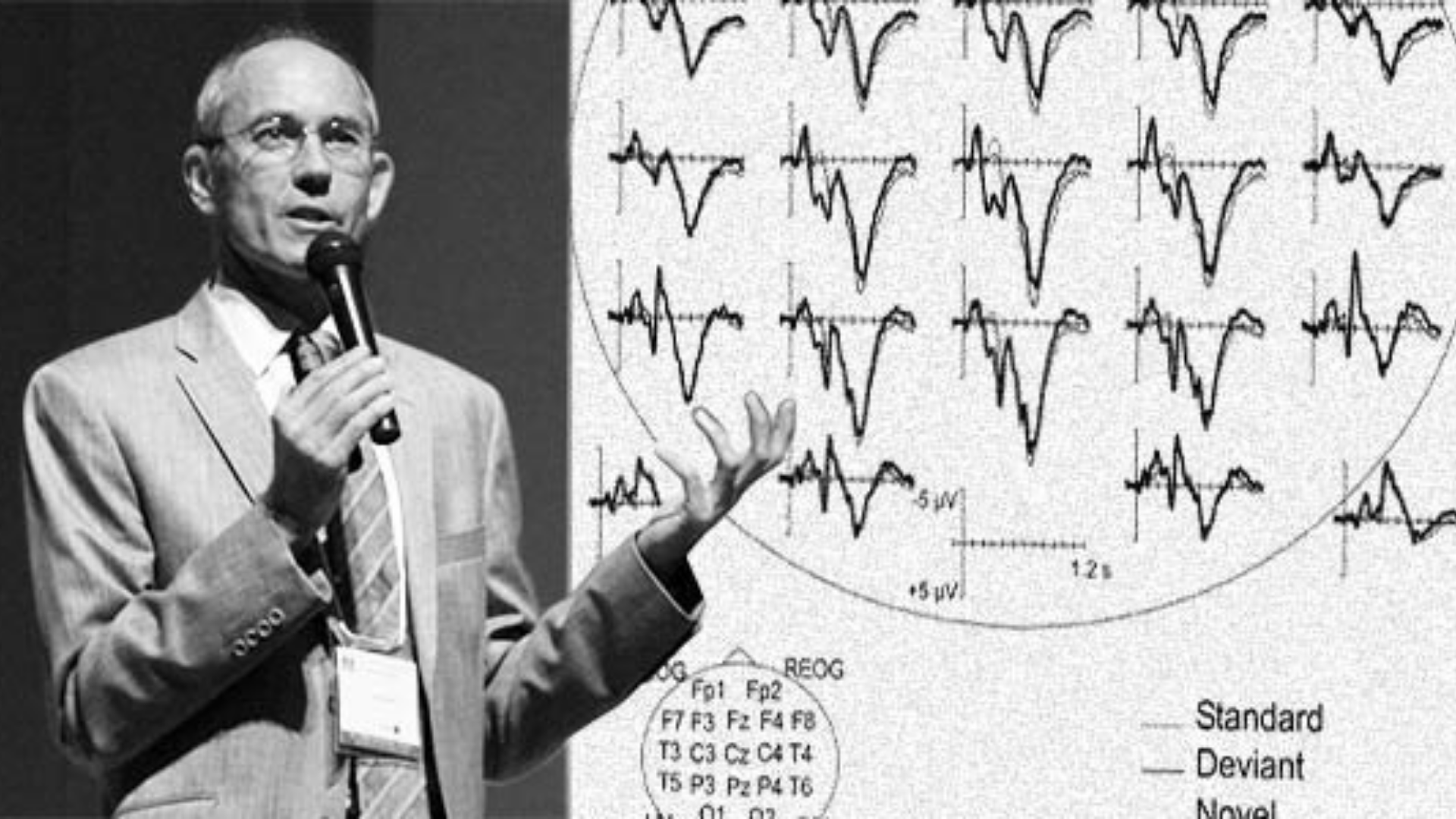
With regard to Forbrain and as a critical factor to be taken into consideration, it has to be highlighted that the cochlea, its basilar membrane, and the encoding of the incoming sound signal into a neural pulse can be considered as a bottleneck through which both air conducted and bone-conducted sounds converge in their way through the central auditory system (Stenfelt et al., 2003; 2005). Hence, if the amplitude and phase of a particular sound are appropriately adjusted, its cochlear signal should be canceled out. This was already anecdotally observed by von Békésy for a 0.4 kHz tone in humans, and subsequently replicated in animals and humans for frequency ranges of 0.1 to 15 kHz (Wever and Lawrence, 1954; Khanna et al., 1976). This is important because, through the dynamic filter implemented in Forbrain, unforeseen and random changes in the bone conduction signal may result in that may eventually cancel out, or at least modify substantially with the air conduction signal to the very same sounds (i.e., the one uttered by the user that reaches the air canal and the emitted by the bone canal after modification by the Forbrain filters resulting in unexpected, rare and potentially surprising changes in the auditory signal that it is finally transduced at the cochlea and conveyed to the central auditory system.
It is proposed here that Forbrain may indeed induce plastic changes in the central nervous system by at least two independent but related neural mechanisms: 1) by challenging the audio-vocal loop through the modified speech signal leading to an enriched acoustic environment yielding auditory plasticity, and 2) by forcing a series of executive mechanisms of attention control to cope with the involuntary attention signals triggered by the mismatching speech inputs. The final outcome of all these processes may be the reinforcement of the executive mechanisms of attentional control, resulting in better concentration, stronger resistance to distracters, improvements in working memory capacity, and the feeling of being more focused.
Compared to many other training and rehabilitation methods for communicative skills, language improvement, and cognitive enhancement, Forbrain has the advantage that although active --it requires the user to speak aloud and to follow a certain regime of exercises-- it imposes very few demands on the user. Indeed, whereas other methods require the attainment of a certain level of performance on a range of exercises of increasing difficulty over several weeks, Forbrain exercises are tailored to the user's motivation, commitment, and willingness to follow the training. This is obviously a strong competitive asset.
Critically, if the "natural" input of the expected voice via the air canal is manipulated by providing "unnatural" (the "extra" bone conduction feedback) and therefore "unexpected" input through the bone canal, the sensory predictions triggered by the efferent copy are violated, resulting in necessary adjustments to the model of the sensory expectation. This template violation by an unexpected input necessarily results in an attentional challenge, as the auditory system identifies the incoming signal as "deviant", "rare" or "unexpected", resulting in an involuntary attention switch (Escera et al., 1998). As a matter of fact, a large body of evidence has indicated that novel or unexpected auditory stimuli trigger an involuntary attention switch toward the incoming stimulus resulting in the behavioral distraction of the ongoing primary task and a concomitant adjustment of brain activity in the underlying neural network (see reviews in Escera et al., 2000; Escera and Corral, 2007). Thus, under conditions of auditory feedback through bone conduction, one might expect to submit the auditory system to a regime of constant challenge through novel or unexpected stimuli, increasing attentional demands. Furthermore, the implementation of the gating filter introduces frequency and temporal manipulations on the emitted voice so that it further deviates the incoming signal from its efferent copy, increasing the attentional challenge driven by Forbrain even more.
The audio-vocal loop
Forbrain's® use of the dynamic filters alters the user's speech signal that is then sent back via a bone conductor and mismatches the corollary discharge (efferent copy) of the planned sounds. This forces online fine-grained adjustments in the audio-vocal loop, leading to auditory plasticity. The vocal (motor) system sends an efferent copy or corollary discharge of the sound it aims at producing so that the encoding of the auditory input resulting from the self-produced sounds is attenuated in the auditory system (Aliu et al., 2008; see Wolpert et al., 1995; Crapse and Sommer, 2008; Scott, 2013).
It is well established that the auditory cortex can undergo plastic changes in response to behaviorally relevant sounds (Fritz et al., 2005; Nelken, 2009), such as those that are conditioned to reward or punishment in animal experiments. What seems to be critical for inducing these plastic changes in the behavioral importance of the stimulus. A paramount example of this property of the auditory system is seen in the brains of musicians, who show major functional and anatomical differences compared to non-musicians (Zatorre, 2013). Yet, auditory plasticity is not only seen in active conditions. For example, Eggermont and colleagues demonstrated in cats that passive long-term exposure to a spectrally enhanced acoustic environment caused a massive reorganization of the tonotopic map in the auditory cortex (Noreña et al., 2006; Pienkowski and Eggermont, 2012). Also, brain plasticity induced by passive music listening (one hour daily during two months, of self-selected materials) was observed in a study of patients recovering from a stroke (Särkämo et al., 2008). These patients showed enhanced recovery in verbal memory, focused attention, and several mood measurements, that were maintained after 6 months of the treatment.
The cerebral network for language includes Broca's area in the frontal lobe of the left hemisphere, and its comprehension counterpart, located in the Wernicke's area in the posterior bank of the superior temporal lobe. There is, however, a particular auditory-related area located in the posterior part of the Planun Temporale of the left hemisphere that is also involved in speech production, the so-called Spt area (Hickok et al., 2000, 2003). Spt is activated during the passive perception of speech and during covert (subvocal) speech articulation (Buschbaum et al, 2001, 2005). It is highly correlated with the pars opercularis (Buschbaum et al., 2005) --an anatomical region subserving part of Broca's area -- with which it is densely interconnected through white matter tracts (Hickok et al., 2011). Hence, by being situated in the middle of a network of auditory (superior temporal sulcus) and motor (pars opercularis) areas, the Spt has been considered as the hub of sensorimotor integration for speech and related vocal-tract functions (Hickok et al., 2010), being implicated in auditory feedback control of speech production. It has been proposed that the Spt region works as a control mechanism for dynamically altering the signals of the planned speech as it is being produced (Hickok et al., 2010).
This is very relevant for Forbrain because the sensory feedback provided by the bone conductor and the dynamic filters do really violate the internal predictions, leading necessarily to online readjustments and retuning of the internal model for speech. For example, a study showed that persistent developmental stutterers display a deficient representation of the sounds of their native language (phonemes), in the presence of preserved acoustic features representation, as measured in passive conditions (Corbera et al., 2005). Moreover, these deficient phoneme representations correlated with the severity of the stuttering, supporting the theoretical model described above. An important implication of the model in general and in particular regarding Forbrain, is that using one's own voice is sufficient to generate corrective signals for motor speech acts, as hearing other speakers' voices is sufficient to learn and tune new motor speech patterns. Hence, one can expect that motor-speech networks in the frontal cortex are activated during passive speech listening, at the same time that a profound system recalibration is taken place online at a very subtle level during normal speech production. Now, if the expected incoming signals from one's own voice are profoundly violated, as occurs with Forbrain, dramatic plastic changes are to be expected in the audio-vocal loop.
Mechanisms of attention control
The unexpected violations of the template predictions of self-emitted sounds trigger a cascade of involuntary attention processes reorienting attention back to the primary task (e.g., the ongoing message) all of which occurs on a sub-second scale. Indeed, a consequence of the mechanisms explained above is that the sensory predictions triggered by the efferent copy of the speech are violated by the manipulated bone conduction input, resulting in necessary adjustments to the model of the sensory expectation. This template violation by an unexpected input results in an attentional challenge, as the auditory system identifies the incoming signal as "disruptive" yielding an involuntary attention switch (Escera et al., 1998). A large body of evidence has indicated that novel or unexpected auditory stimuli trigger an involuntary attention switch toward the incoming stimulus resulting in the behavioral distraction of the ongoing primary task and a concomitant adjustment of brain activity in the underlying neural network (see reviews in Escera et al., 2000; Escera and Corral, 2007). The activation of this neural network can be tracked by the recording of its neural signature at the scalp, that is to say, by extracting the corresponding event-related brain potentials from the ongoing EEG. A series of waveforms have been identified that conform to the so-called distraction potential (DP; see Figure 1), and that reflect three successive stages in the involuntary attention chain: the mismatch negativity (MMN) reflecting detection of the disparity (Escera et al., 1998); the novelty-P3, reflecting the effective orienting of attention towards the eliciting sound (Escera et al., 1998), and reorienting negativity (RON) (Schröger and Wolff, 1998; Escera et al., 2001). Also, the DP can reveal the interactions between top-down and bottom-up forms of attention, such as during working memory load (San Miguel et al., 2008) and emotional challenge (Domínguez-Borràs et al., 2009). As discussed above, it is very likely that by the nature of the manipulations it introduces to the voice of users, Forbrain induces a remarkable challenge to this cerebral network for involuntary attention control, and the recording of these involuntary attention-related potentials provides an unprecedented framework to validate its principles of action.
Moreover, the auditory system can also cope with distracters, easily reorienting attention back to task performance after a transitory attention switch (Escera et al., 2001; Schröger and Wolff, 1998; see Escera and Corral, 2007). The final outcome of all these back and forth processes of orienting and reorienting of attention might be a general improvement in attention control capabilities, allowing better protection against distracters and a better
